The Korean text of this paper can be translated into multiple languages on the website of http://jksee.or.kr through Google Translator.
정안천에서 어류 군집 기반의 생태학적 연결성 분석
Abstract
This study focused on the longitudinal connectivity analysis, based on fish communities in the aspect of spatial variations of 34 sites from upstream to downstream. The Fish Assessment Index (FAI), based on ecological guilds and species composition, was applied to compare the ecological health among the sampling sites. The total number of 35 fish species, were sampled during the study period with dominant species of Zacco platypus (35.4%) and Zacco koreanus (11.8%). Cluster analysis showed that all sites were categorized as 4 distinct group communities (A, B, C, D). Non-metric multidimensional scaling analysis showed distinct difference between upstream (A, B) and downstream groups (C, D). Indicator Value Analysis (IndVal method), conducted to identify indicator species within each group, indicated that the species favoring upstream environments Rhynchocypris oxycephalus for the Group A and Iksookimia koreensis for the Group B. In contrast, the analysis indicated that the species with a preference for downstream habitats Pseudogobio esocinus for the Group C and Carassius auratus for the Group D. Ecological health, based on the FAI scores, showed a declining pattern toward the downstream, along with an increasing proportion of tolerant species and omnivores within the fish community. Overall these findings suggest the compositions of fish communities, as four groups, were modified by the weir structures and organic matter, nutrient pollutions.
Key words: Community ecology, Fish assemblage, Longitudinal connectivity, Ecological health assessment
요약
본 연구에서는 정안천 상류부터 하류까지의 지점 별 어류 군집 변화를 분석하였다. 2021년, 정안천에서 34지점을 선정하여 어류를 채집, 종 조성 자료를 얻었다. 이를 토대로 군집 내 어류 길드, 종 조성에 기초한 어류 평가 지수(Fish Assessment Index, FAI)을 적용, 각 지점의 생태적 건강도를 비교하였다. 정안천 34개 지점에서 채집된 어류는 총 35종, 7,436개체로, 우점종은 피라미(2,634개체, 35.4%), 아우점종은 참갈겨니(877개체, 11.8%)로 확인되었다. 각 지점 사이의 유사성을 비교하기 위해 군집분석(Cluster analysis)을 수행한 결과, 각 지점은 총 4개의 집단(A, B, C, D)으로 구분되었다. 이후 비모수적다차원척도법(Nom-metric multidimentional scaling)을 적용한 결과, 상류에 위치한 지점들(A, B)이 높은 연속성, 유사도를 보이며 하류의 지점들(C, D) 또한 서로 유사하게 나타난 한편 상류와 하류를 비교하였을 경우에는 차이를 확인할 수 있었다. 각 군집에 대해 지표종 수치(IndVal method)를 산출하였을 때, 상류의 2개 군집에서는 버들치, 참종개 등 상류에 주로 서식하는 어종이 지표종으로 나타난 반면 하류 군집에서는 모래무지, 붕어와 같은 하천 중류, 하류를 선호하는 어종이 높은 지표종 수치를 보였다. 또한 FAI를 통해 지점별 생태적 건강도를 비교하였을 때, 하류 군집에서 낮은 수치를 확인할 수 있었다. 상기 결과를 종합하였을 때, 상류와 하류 군집이 구분되는 지점(S14 ~ 15) 이후의 하류 지점에서 인간의 교란에 따라 생태적 변화가 유발됨을 판단할 수 있었다.
주제어: 군집생태학, 어류 군집, 종적 연결성, 생태학적 건강성 평가
1. 서 론
하천은 풍부한 생물다양성을 지니고 있으며 사회에 필수적인 생태계 서비스를 제공한다[ 1, 2]. 하천은 수문학적 연속성을 지니며, 각 생태계 요소 사이에서 에너지, 물질, 개체를 수송하는 통로 역할을 한다[ 3]. 이러한 하천의 연속성은 하천 생태계의 기능을 유지하기 위해 필수적인 요소로 하천의 고유한 특성이라고 할 수 있다[ 4]. 종방향의 연속성을 갖는 자연상태의 하천에서 생물의 분포 또한 연속적인 변화를 나타내며 이는 하천 연속성 개념(River Continuum Concept)으로 설명된다[ 5]. 하천 연속성 개념 하에서 상류부터 하류에 이르기까지 하천의 유기물 조성, 하천의 규모, 유속 등 환경 요소가 변화하며 생물 군집 또한 이러한 변화와 수반하여 분포하게 된다. 하지만 인간 활동에 의해 하천 및 수역에 교란이 발생할 경우, 하천 환경은 자연상태에서 예상되는 종적인 변화와 다른 양상을 보인다. 하천 인근에 도심지, 농경지가 형성될 때, 특정 지점에서 유기물, 영양염류 등이 추가로 유입되며, 이는 지리적 구배를 따르지 않는 이질적인 환경을 야기한다[ 6, 7]. 이러한 교란은 하천에 서식하며 연속적으로 변화하는 생물상 분포에 영향을 미치게 된다. 농업의 영향으로 식생이 감소하고 비료에 의한 영양염류 농도가 높아지는 경우, 상류와 하류 전체에 걸쳐 오염에 내성도가 높고 유사한 저서무척추동물 군집이 형성될 수 있다[ 8]. 또한, 하천 상류 지역이 도시화로 인한 오염에 노출되었을 때, 해당 지역에서 어류 군집에 내성도가 높은 외래종 비율이 높아지는 동시에 어류의 개체수가 감소함으로써 하천에 불연속적인 군집 분포가 나타나는 현상이 보고된 바 있다[ 9]. 한편, 댐, 보 등의 구조물이 건설되면서 하천의 물리적 연결성이 단절되어 서식지가 단편화되며 수생태계가 악화되는 문제가 발생하고 있다[ 10, 11]. 인공 구조물의 건설은 연속성의 단절을 유발하는 동시에 하천의 유속, 유량, 탁도 등의 물리적 환경과 화학적 수질 및 생태계에 영향을 끼치게 되며 하천 내 생물 다양성에 변화를 불러온다[ 12]. 특히, 수생태계 환경과 밀접한 관계를 갖는 어류의 경우 이러한 교란에 직접적으로 영향을 받게 된다. 인공 구조물은 어류 산란에 필수적인 서식지 이동을 막아 어류 개체군을 감소시키며 이와 같은 지리적 격리는 유전적 고립과 지역적 멸종의 원인이 될 수 있다[ 13, 14]. 또한 인공 보는 물의 흐름을 막아 하천에 유속이 느리고 수심이 깊은 소를 형성, 이에 따라 어류 군집에서 유수성 어종이 줄어들고 잡식성, 정수성 어종 비율이 높아지게 된다[ 15]. 이러한 인공 구조물에 의한 서식지의 질적 하락은 장애물의 규모가 작더라도 발생할 수 있으며 어류 군집 분포에 유의한 영향을 미치게 된다[ 16, 17]. 우리나라의 경우 지난 수십년간 급격한 산업화, 도시화를 거쳐왔으며 이로 인해 하천의 수질 오염, 서식지 파괴와 같은 다양한 문제가 발생하였다[ 18, 19]. 이에 따라 하천 생태계에 서식하는 생물 군집 내 내성종 비율이 높아지고 다양성이 하락하는 등 수생태계 건강성의 악화가 보고된 바 있다[ 20, 21]. 또한 몬순기후의 영향으로 강우가 연중 특정 시기에 집중됨에 따라, 안정적인 수자원 확보를 위해 전국에 많은 인공 보 및 댐이 건설되었다. 해양수산부의 국가어도정보시스템에 따르면 하천에 설치된 보 개수는 2022년 기준 33,914개에 달한다. 이에 따라 많은 하천 서식지가 변형되었으며 개체의 이동 차단 및 하천의 유속, 하상 구조의 변화로 인한 어류 군집 변화가 보고된 바 있다[ 22, 23]. 이러한 인공 구조물의 영향은 하천의 수 문학적 특성, 서식하는 생물 등 요인에 따라 다양한 형태로 나타나기 때문에 하천 특성에 기반한 평가 및 복원 방안이 요구된다[ 24, 25]. 현재 환경부에서 관계 부처의 도움을 받아 수생태계 연속성 조사를 수행하고 연속성 단절에 대한 기준을 마련하기 위해 2017년 물환경보전법 제22조의2가 신설되었다. 하지만 아직 국내의 수생태계 연속성의 단절 및 훼손에 대해 제시된 사례가 부족한 실정으로 실제 어류의 이동성 평가, 채집을 통한 모니터링, 구조물 특성 분석 등 다양한 연구가 필요하다[ 26]. 본 연구에서는 정안천 상류에서 하류지역까지의 어류 군집을 채집하고 분석을 수행하여 하천의 수생태계 연속성 및 건강성을 확인하였다. 통계적 기법을 활용하여 상류부터 하류에 이르기까지의 종 조성 변화를 분석하였으며 어류 군집의 생태적 길드 분석 및 건강도 평가를 통해 각 지점의 수생태계 건강성을 비교하였다. 실제 하천 생태계와 정안천 환경에 대한 정보를 제공함으로써 향후 생태계 복원 및 관리의 기초 자료를 제공하고자 하였다.
2. 재료 및 방법
2.1. 조사 지점
조사 지역은 충청남도 공주시에 소재한 유로연장 29.6 km, 유역 면적 161.7 km2의 정안천으로, 정안천은 정안면에서 발원하여 금강으로 합류되는 금강의 지류하천이다. 본 연구에서는 정안천에서 인위적인 교란에 의한 어류 군집의 연속성과 단절을 분석하기 위해 상류부터 하류까지 총 34개의 조사 지점을 선정, 어류를 채집하였다( Fig. 1, Table 1). 34개 지점은 인공 구조물에 의한 영향을 파악하기 위해 국가어도정보시스템을 참조하여 인공 보가 존재하는 곳을 조사 지점으로 선정한 결과이다. 각 보는 공주시 또는 한국농어촌공사의 관리 하에 있으며, 보의 규모는 폭 0.45~2.00 m, 길이 11~170 m, 높이 0.30~1.51 m의 범위로, 하천 하류에서 하천의 규모가 확대됨에 따라 대체로 하류의 보가 규모가 커지는 경향을 보인다. 하천의 연속성 단절을 완화하기 위한 어도는 34개 중 11개의 보에 설치되어 있다. 한편 인근의 오염원으로 농업지역을 관통하는 하천의 특성상 하류 지점 인근에 경작지가 다수 존재하며, 사현천이 합류되는 광정리 인근에는 2개의 농공단지가 위치하여 하수처리수가 방류되는 특징을 가진다.
2.2. 어류 현장 채집
어류 채집은 환경부의 수생태계 연속성 조사 및 평가 방법 등에 관한 지침[ 27]에 의거하여 수행되었다. 각 지점에 존재하는 보의 하류부에서 조사를 수행하였으며, 조사 시기는 2021년으로 환경이 안정된 봄철 및 가을철에 각 지점 2회 조사하였다. 조사 도구는 투망(망목 7×7 mm) 및 족대(망목 5×5 mm)를 사용하였으며 정량적인 채집을 위해 투망 10회, 족대 30분의 일정한 횟수 및 시간을 준수하였다. 조사 지점의 서식지 특성을 반영할 수 있도록 여울, 소 등 지점에 존재하는 다양한 환경을 포함하여 채집을 진행하였다. 채집된 어류는 현장에서 즉시 분류, 동정하여 개체수를 확인한 후 현장 방류하였으며 일부 현장 동정이 어려운 경우 10% 포르말린 용액에 고정하여 실험실에 운반 후 동정하였다.
2.3. 내성도 및 섭식 길드 분석
각 지점의 어류 군집 조성의 특성을 분석하기 위해 지점별로 채집된 어류는 Ohio EPA [ 28]와 Barbour 등[ 29]의 방법에 기초하여 내성도 및 섭식 길드에 따라 분류하였다. 내성도 길드는 수질 오염 등 환경 교란에 취약한 민감종(Sensitive species, SS), 오염된 환경에 내성이 강한 내성종(Tolerant species, TS), 민감종과 내성종 사이의 내성도를 보이는 중간종(Intermediate species, IS)으로 분류되었다. 섭식 길드는 어류가 먹이원으로 이용하는 자원에 따라 육식종(Carnivores, C), 충식종(Insectivores, I), 초식종(Herbivores, H), 잡식종(Omnivores, O)로 분류되었다. 육식종은 어류 등을 섭식하며 충식종은 주로 수서 곤충을 섭식한다. 초식종은 부착조류와 플랑크톤을 섭식하며 잡식종은 동물성 및 식물성 먹이원 모두를 이용한다.
2.4. 어류 평가 지수(Fish Assessment Index, FAI)를 적용한 생태계 건강성 평가
어류는 수생태계의 일원으로 다양한 생태적 지위의 종을 포함하고 있으며, 물리적, 화학적, 생물적 환경 요소에 대한 종합적인 평가 지표로 사용될 수 있다[ 30]. 실제 미국을 비롯한 다양한 국가에서 어류 종 조성에 기반한 생태계 건강성 평가 기법이 확립되어 있으며 국내에서도 이용된 바 있다[ 19, 28]. 본 연구에서는 상류부터 하류까지의 생태계 건강성 변화를 파악하기 위해 환경부의 지침[ 31]에 의거하여 어류 종 조성에 기반한 건강성 평가 모델(FAI)을 적용, 8개 메트릭 항목에 기반한 점수를 산출, 비교하였다. 8개의 메트릭 항목은 M1(국내종의 총 종수), M2(여울성 저서종수), M3(민감종수), M4(내성종의 개체수 비율), M5(잡식종의 개체수 비율), M6(국내종의 충식 종 개체수 비율), M7(국내종의 총 개체수), M8(비정상종 개체수 비율)로 구성되었으며, 환경부의 기준에 의거하여 각 메트릭에 ’12.5’, ‘6.25’, ‘0’의 점수를 부여하고 8개 매트릭 항목의 합계로 FAI score를 계산하였다. 합산된 FAI score는 총점 100점으로 0~20을 ‘매우 나쁨’, 20~40을 ‘나쁨’, 40~60을 ‘보통’, 60~80을 ‘좋음’, 80~100을 ‘매우 좋음’으로 등급을 구분하여 서식지 건강성의 척도로 사용하였다.
2.5. 통계적 기법울 이용한 군집 유사도 및 지표종 분석
지점별 어류 군집의 유사도와 변화 정도에 따라 지점을 분류하기 위해 군집분석(Clustering analysis)을 수행하였다. 각 지점에서 2회 채집된 어종 개체수 데이터를 합산한 지점별 어류 군집 데이터를 통계 소프트웨어 R (version 4.3.2) [ 32]의 패키지 ‘vegan’ [ 33]을 사용하여, chord 변환을 적용하였으며 유클리드 거리(Euclidean distance)에 따른 유사도를 구하였다[ 34]. 이후 지점별 유사도를 바탕으로 각 조사지점간 유사도를 Ward의 방법으로 분류[ 35], 덴드로그램(Dendrogram)으로 시각화하였다. 이와 함께 비모수적다차원척도법(Non-metric multidimensional scaling, NMDS)을 사용, 지점별 유사도를 시각적으로 표현하였다[ 36]. NMDS분석에서는 R의 패키지 vegan [ 33]의 metaMDS 함수를 이용, 브레이커티스(Bray-Curtis) 방법[ 37]에 따라 지점별 종 조성의 유사도를 산출하였다. 군집분석에서 정의된 각 집단의 특성을 대표하는 지표종을 확인하기 위해 ‘labdsv’ [ 38] 패키지를 사용, 지표종 수치 분석(Indicator value analysis)을 수행하였다[ 39]. 집단에 속한 각 종들의 지표종 수치(IndVal)는 집단내에서의 특정 종의 평균 풍부도와 전체 지점의 해당 종 풍부도 비율(Specificity)과 집단 내에서 종이 발견되는 지점의 비율(Fidelity)의 곱으로 계산되었다[ 34]. Specificity는 종이 특정 집단에만 존재할 경우 최댓값을 보이며, Fidelity는 해당 종이 집단에 속하는 모든 지점에서 발견될 경우 최댓값을 보인다. 도출된 지표종 수치는 10000회의 순열검정(Permutation test)를 통하여 유의수준 5% 수준에서 유의성을 검정하였다[ 34].
3. 결과 및 고찰
3.1. 정안천의 어류상
조사기간 중 34개 지점에서 총 35종 7,436 개체의 어류가 채집되었다( Table 1). 출현 종 중 우점종 및 아우점종은 우리나라 하천 상류 및 중류에서 흔히 발견되는 피라미와 참갈겨니로 나타났으며 2,634개체(35.4%), 877개체(11.8%) 채집되었다. 피라미의 경우 항존도(Constancy) 100%로 상류부터 하류에 이르기까지 모든 지점에서 출현하는 동시에 대부분의 지점에서 우점종으로 확인되었다. 하지만 참갈겨니는 항존도 73.5%의 비교적 낮은 수치를 나타냈으며 상류에서는 높은 우점도를 보였으나 하류에서는 출현하지 않거나 소수의 개체만 확인되었다. 항존도의 경우 참갈겨니보다 돌고기에서 더욱 높은 수치를 보였는데, 이러한 결과는 피라미, 돌고기가 비교적 넓은 서식지 범위를 지니는 것에 반해 수온이 낮고 유속이 빠른 상류 환경을 선호하는 참갈겨니의 특성에 의한 것으로 사료된다. 참갈겨니와 피라미 다음으로는 붕어가 645개체(8.7%)로 비교적 높은 상대 풍부도를 보였다. 그러나 붕어는 47.1%의 낮은 항존도를 보였으며 이는 정수역을 선호하는 붕어의 특성상 붕어가 하천 상류 보다는 하류 구간에서 주로 출현한 것이 그 원인으로 사료된다. 전체 지점에서 법정보호종은 출현하지 않았으며 고유종은 참종개 등 총 11종이 출현하였다. 외래종은 배스, 블루길 2종이 출현하였으며 하천 하류 지점에서 확인되었다.
3.2. 지점별 군집분석 및 NMDS분석 결과
지점별 어류상 자료를 바탕으로 수행된 군집분석 결과를 덴드로그램으로 시각화 하였을 때 하천 상류부터 하류까지의 하천 어류상은 총 4개의 집단(Group A, B, C, D 이후 G-A, G-B, G-C, G-D로 사용)으로 구분되었다( Fig. 2). 먼저 G-A의 경우 S1에서 S4까지의 4개의 최상류 지점으로 구성되었다. G-A에 속한 지점에서 8종 472개체의 어류가 채집되었으며, 가장 높은 우점도를 보인 종은 참갈겨니(59.8%)로 G-A에 속한 지점전체에서 우점종으로 나타났다. 참갈겨니 다음으로는 버들치(29.0%)가 높은 상대풍부도를 보였다. G-B는 S5에서 S14까지의 상류 지점, 그리고 S22 총 11개 지점으로 이루어졌다. G-B의 경우 19종 1,900개체의 어류가 채집되어 피라미가 44.5%의 상대풍부도로 우점종, 참갈겨니가 26.3%의 상대풍부도로 아우점종으로 나타났다. G-C는 하천 중류 및 하류에 위치한 S15, S18~21, S23~26, S31~34의 13개 지점으로 구성되었다. G-C에서는 34종 3,279개체가 채집된 한편 우점종은 G-B와 동일하게 피라미로 집단 내에서 43.5%의 상대풍부도를 보였으며, 아우점종은 납자루(12.1%)로 확인되었다. G-D는 하천 하류 지점인 S16~17, S27~30 6개의 지점으로 구성되었다. G-D 지점에서는 34종 1,785 개체가 채집되었으며, 우점종은 붕어로, 29.6%의 상대풍부도를, 아우점종은 피라미로 19.0%의 상대풍부도를 보였다. NMDS분석을 수행, 군집분석 결과를 반영하여 지점 사이의 종 조성 차이를 평면에 시각화한 결과( Fig. 3), 최상류에 위치한 G-A와 이에 이어지는 상류의 G-B가 평면상에서 서로 인접하게 배치되었으며 하류에 위치한 G-C와 G-D 또한 서로 비교적 높은 유사도를 보였다. 하지만 상류의 집단과 하류의 집단을 비교하였을 떄, 차이가 나타남을 확인하였다. 이는 G-B와 G-C의 경계가 되는 S14와 S15 사이에서 생물상의 변화가 존재함을 나타내며 이를 기준으로 하천의 어류 군집과 생태적 특성이 변화함을 의미한다. 한편, S22는 하류 지점에 위치하였으나 예외적으로 G-B에 속하였는데, NMDS분석 상에서는 집단 G-C에 속한 지점들과 보다 높은 유사성을 보였다. 해당 지점에서는 상하류 지점들(S21, S23)과 달리 주로 상류에서 서식하는 참갈겨니가 다수 출현하였으며 이는 인근에서 소하천의 영향으로 사료되며 실제로 납자루 등 근접한 지점에서 주로 발견되는 어종들 또한 채집되어 하류 지점의 특징을 함께 나타냈다. 군집 분석과 NMDS분석 결과를 종합하였을 때, 어류 군집은 상류부터 하류에 이르기까지 전반적으로 연속해서 변화하는 한편, 특정 지점에서 종 조성의 변화가 급격히 나타나는 것으로 사료된다.
3.3. 지표종 분석
각 집단에 대한 지표종 수치 분석 결과는 다음과 같다( Table 2). 상류 지점으로 이루어진 G-A와 G-B에서는 각각 버들치(IndVal = 0.840, p < 0.001)와 참종개(IndVal = 0.777, p < 0.001)가 가장 높은 IndVal 수치를 보이는 지표종으로 확인되었다. 한편 G-C에서는 모래무지(IndVal = 0.745, p < 0.001), G-D에서는 붕어(IndVal = 0.907, p < 0.001)가 가장 높은 수치를 보였다. 버들치와 참종개의 경우 유량이 적고 수온이 낮으며 여울이 발달한 하천 상류에 주로 서식하는 어류인 반면 모래무지, 붕어는 유속이 비교적 느린 중류 및 하류에 주로 서식하는 어류로써 상류의 집단에 속한 지점들과 하류의 지점 사이에서 생태적 특성, 종 조성이 달라짐을 나타낸다. 모래무지는 하상에 모래가 많은 곳에서 주로 출현하는 종으로 이에 따라 유속이 빨라 하상에 모래가 유지되기 어려운 환경보다 비교적 유속이 느린 환경을 선호한다[ 40]. 또한 붕어의 경우 정수성 어류이자 내성종, 잡식성 어종으로 분류되는 특성을 가지며 보의 영향과 기타 인위적 원인으로 하천생태계에 교란이 존재한 가능성을 시사한다[ 15, 30]. 따라서 G-C와 G-D에 속한 지점에서 상류 지점에 비해 급격한 유속의 감소 및 서식지 훼손이 발생한 것으로 사료된다.
3.4. 어류 군집의 생태학적 길드 및 건강성 분석
각 지점별 어류 군집의 내성도, 섭식 길드를 분석하여 군집분석에서 도출된 4개 집단의 특성을 비교하고자 하였다( Fig. 4). 내성도 길드를 비교하였을 때, 상류의 G-A와 G-B의 경우에는 내성종의 비율이 낮고 대부분의 어류가 민감종 및 중간종으로 구성된 반면, 하류 지점에서는 내성종이 다수 출현하였다. 이러한 결과는 하류 지점에서 수환경이 악화됨을 나타내며, 인근에 위치한 농경지, 하수처리시설 등의 영향을 받고 있음을 추측할 수 있다. 또한 섭식 길드의 경우 상류의 집단에서는 충식 종 비율이 높게 나타났으나 하류로 갈수록 잡식종의 비율이 증가하는 경향을 보였다. 이러한 충식종의 출현은 하천의 서식 환경이 악화될 경우 감소하며 하천 환경을 간접적으로 나타내는 지표로 이용될 수 있다[ 41]. 따라서 생태적 길드 분석 결과를 종합하였을 때, 하천 하류 지점들, 특히 Group D에 해당하는 지점들에서 서식지 교란의 위험성이 높게 나타난다. 8개 항목의 FAI를 통해 각 지점의 생태학적 건강성을 분석한 결과는 다음과 같다( Table 3, Fig. 4). G-A는 평균 84.4(매우 좋음), G-B는 평균 77.3(좋음), G-C는 평균 61.1(좋음), G-D는 평균 45.8(나쁨)의 FAI를 보였다. 상류의 G-A와 G-B에서는 14개 지점 중 FAI 총합 80점 이상으로 매우 좋음에 속하는 지점이 10개 지점으로 전반적으로 건강성이 우수한 것으로 나타났으나, 하류에 위치한 G-C, G-D의 경우 20개 지점 중 13개 지점이 FAI 총합 60점 이하로 보통 이하의 수준으로 분류되었다. 하류 지점에서 높은 잡식종, 내성종 비율과 낮은 FAI score를 나타내는 것은 오염원 및 기타 교란에 의한 수생태계 악화를 시사한다. 실제 환경부( www.me.go.kr)에서 제공하는 전국 공공폐수처리시설 운영현황에 따르면 S15인근에 위치한 정안농공단지와 보물농공단지에서 2021년 기준 일 평균 279 m 3, 74 m 3의 방류수를 각 농공단지에 위치한 하수처리장에서 방류하였다. 특히 정안농공단지의 경우 방류수의 평균 총 질소 농도가 약 12 mg/L에 달하며 이러한 질소의 유입은 하천 수생태에 영향을 미칠 수 있다[ 42]. 또한 정안천 하류 지점에는 농경지가 존재하기 때문에 비점오염원에 의한 교란에도 노출되어있는 상황이다. 이를 고려하였을 때, 정안천 하류 지점에서 교란에 의한 영향이 어류 군집에서 나타나는 것으로 사료되며 상류 지점과의 종 조성 차이를 유발한 것으로 판단된다 .
4. 결 론
본 연구에서는 정안천에 존재하는 34개 보의 하류 지점에서 어류를 채집, 종 조성 자료를 분석하여 하천 상류부터 하류까지의 생물상, 생태 변화를 확인하고자 하였다. 연구 결과, 정안천 상류부터 하류까지의 하천 생태계는 지리적 구배를 따라 변화하는 한편, 인간의 교란 또한 영향을 미쳐 특정 지점을 경계로 상류 지점들과 하류 지점 사이에 이질성이 야기되는 것으로 분석된다. 군집분석에서 각 지점은 총 4개의 집단으로 분류할 수 있으며 각 집단의 위치는 최상류, 상류, 그리고 하류로 구분할 수 있었다. 지점의 종 조성에서 S14와 S15를 경계로 상류에 속한 G-A와 G-B가 서로 높은 유사도를 나타냈으며 하류의 G-C와 G-D가 서로 높은 유사도를 보였다. 하지만 상류와(G-A, G-B)와 하류(G-C, G-D)를 비교할 경우 상하류 사이에서는 낮은 유사도를 보여 상류 14개 지점과 하류 20개 지점 사이에 급격한 종의 변화가 존재함을 확인할 수 있었다. S1에서 S14까지 정안천 상류에 속하는 지점에서는 참갈겨니, 버들치 등 수온이 낮고 여울이 조성된 상류에 서식하는 어류가 높은 비율로 확인되었으나 S15부터 하천 최하류에 이르기까지 모래무지, 붕어 등 중류, 하류에 주로 서식하는 어류가 출현, 높은 개체수를 보였다. 또한 해당 지점을 경계로 하류 지점에서는 내성종과 잡식종의 급격히 비율이 높아지는 동시에 환경 악화에 취약한 충식종과 민감종의 비율이 낮아지는 경향을 보였다. 이에 따라 생태적 건강도를 나타내는 FAI 값이 낮아졌으며 이러한 결과는 하류 S14와 S15 인근과 하류 지점에 존재하는 어류 군집이 인위적인 교란요인에 의해 부정적인 영향을 받음에 따라 상류와 종 조성에서 차이를 나타냄을 시사한다. 이를 고려하였을 때, 향후 정안천의 복원 및 관리를 위해서는 해당 지점을 중점적으로 이화학적 평가 및 물리적 평가를 수행함으로써 수생태계 회복에 기여할 수 있을 것으로 사료된다.
Notes
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Fig. 1.
The dendrogram and map for the cluster analysis based on the fish assemblages collected at the study sites in the Jeongan Stream (red: Group A (G-A), green: Group B (G-B), blue: Group C (G-C), purple: Group D (G-D)) 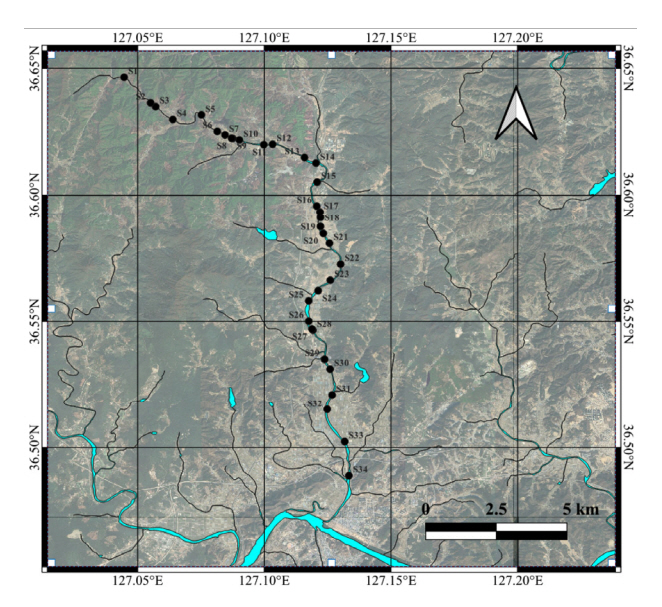
Fig. 2.
Ordination biplot analysis to display the arrangement of study sites using non-metric multidimensional scaling (NMDS), which was based on fish assemblages. The stress value of the NMDS indicates the quality of fit. 
Fig. 3.
Relative abundance of tolerant guilds (a) and trophic guilds (b) in the group of study sites. 
Fig. 4.
The FAI (Fish Assessment Index) score of each study site based on fish communities. 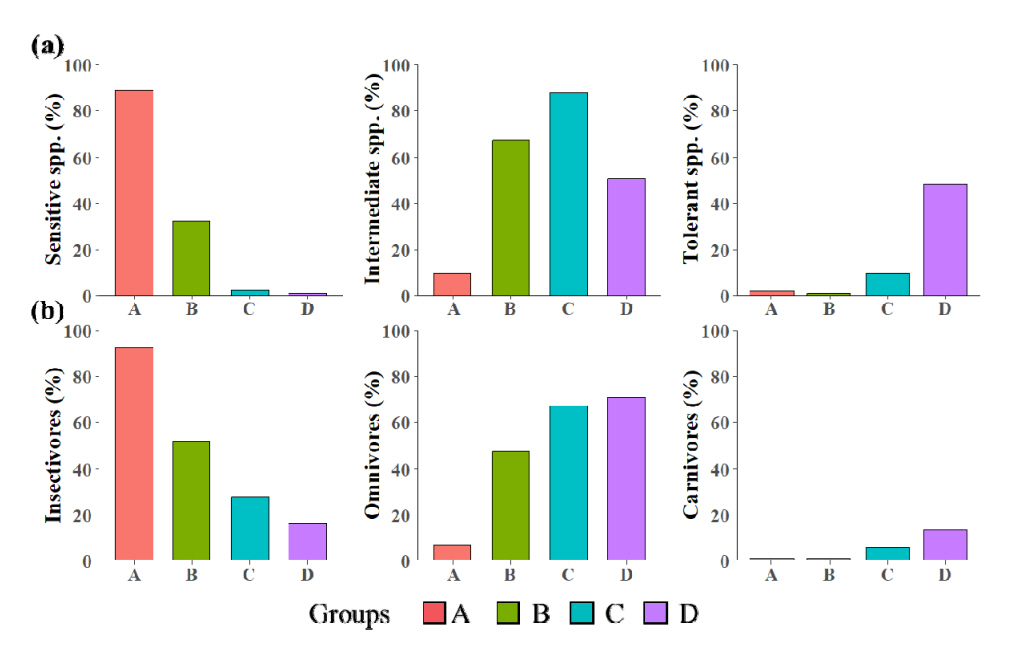
Table 1.
The location of the sampling sites and the weir structures in Jeongan Stream.
|
Sites |
Coordination |
Weir width (m) |
Weir length (m) |
Weir height (m) |
Number of fishways |
|
S1 |
36° 38′ 47.4″N, 127° 02′ 41.1″E |
1.15 |
12 |
0.65 |
0 |
|
S2 |
36° 38′ 11.1″N, 127° 03′ 18.7″E |
1.17 |
11 |
0.63 |
0 |
|
S3 |
36° 38′ 05.8″N, 127° 03′ 25.8″E |
1.25 |
15 |
1.25 |
0 |
|
S4 |
36° 37′ 47.5″N, 127° 03′ 50.3″E |
1.15 |
20 |
0.62 |
0 |
|
S5 |
36° 37′ 54.1″N, 127° 04′ 30.9″E |
0.97 |
25 |
0.63 |
0 |
|
S6 |
36° 37′ 30.7″N, 127° 04′ 53.4″E |
1.10 |
37 |
0.99 |
0 |
|
S7 |
36° 37′ 25.6″N, 127° 05′ 04.5″E |
1.26 |
23 |
0.95 |
0 |
|
S8 |
36° 37′ 20.9″N, 127° 05′ 13.8″E |
1.00 |
28 |
0.70 |
1 |
|
S9 |
36° 37′ 20.7″N, 127° 05′ 15.4″E |
1.00 |
28 |
0.70 |
0 |
|
S10 |
36° 37′ 18.5″N, 127° 05′ 24.8″E |
1.00 |
29 |
0.88 |
1 |
|
S11 |
36° 37′ 11.8″N, 127° 05′ 59.6″E |
1.30 |
52 |
1.51 |
0 |
|
S12 |
36° 37′ 12.4″N, 127° 06′ 12.1″E |
1.30 |
52 |
1.51 |
2 |
|
S13 |
36° 36′ 53.6″N, 127° 06′ 57.4″E |
1.30 |
27 |
1.00 |
0 |
|
S14 |
36° 36′ 45.7″N, 127° 07′ 13.7″E |
1.30 |
66 |
1.24 |
3 |
|
S15 |
36° 36′ 18.7″N, 127° 07′ 15.3″E |
1.17 |
91 |
0.75 |
0 |
|
S16 |
36° 35′ 44.5″N, 127° 07′ 14.8″E |
0.45 |
88 |
1.40 |
1 |
|
S17 |
36° 35′ 36.0″N, 127° 07′ 19.5″E |
0.85 |
130 |
1.00 |
1 |
|
S18 |
36° 35′ 27.9″N, 127° 07′ 20.1″E |
1.20 |
98 |
1.20 |
1 |
|
S19 |
36° 35′ 14.7″N, 127° 07′ 20.1″E |
1.00 |
119 |
0.70 |
0 |
|
S20 |
36° 35′ 04.8″N, 127° 07′ 24.1″E |
1.00 |
78 |
0.80 |
0 |
|
S21 |
36° 34′ 50.8″N, 127° 07′ 32.9″E |
1.00 |
87 |
0.72 |
1 |
|
S22 |
36° 34′ 21.0″N, 127° 07′ 48.7″E |
0.70 |
87 |
1.00 |
1 |
|
S23 |
36° 33′ 58.3″N, 127° 07′ 34.1″E |
0.85 |
90 |
0.50 |
0 |
|
S24 |
36° 33′ 43.4″N, 127° 07′ 16.8″E |
1.00 |
85 |
0.30 |
0 |
|
S25 |
36° 33′ 29.0″N, 127° 07′ 03.3″E |
1.10 |
110 |
1.11 |
0 |
|
S26 |
36° 33′ 00.3″N, 127° 07′ 03.4″E |
1.20 |
100 |
1.40 |
0 |
|
S27 |
36° 32′ 49.1″N, 127° 07′ 08.1″E |
1.20 |
94 |
1.10 |
0 |
|
S28 |
36° 32′ 47.5″N, 127° 07′ 09.3″E |
2.00 |
88 |
0.50 |
0 |
|
S29 |
36° 32′ 06.2″N, 127° 07′ 25.9″E |
0.90 |
110 |
1.40 |
0 |
|
S30 |
36° 31′ 52.1″N, 127° 07′ 33.8″E |
1.10 |
106 |
1.40 |
0 |
|
S31 |
36° 31′ 15.4″N, 127° 07′ 36.7″E |
1.10 |
109 |
0.60 |
0 |
|
S32 |
36° 30′ 54.3″N, 127° 07′ 29.7″E |
1.15 |
170 |
0.70 |
1 |
|
S33 |
36° 30′ 08.3″N, 127° 07′ 54.5″E |
1.00 |
48 |
1.30 |
0 |
|
S34 |
36° 29′ 20.1″N, 127° 08′ 00.5″E |
1.15 |
50 |
0.60 |
1 |
Table 2.
Fish species and individuals collected from the sites in the Jeongan Stream in 2021 (RA: Relative abundance, Cons: Constancy).
|
Scientific name |
Sites
|
Total |
RA (%) |
Cons (%) |
|
S1 |
S2 |
S3 |
S4 |
S5 |
S6 |
S7 |
S8 |
S9 |
S10 |
S11 |
S12 |
S13 |
S14 |
S15 |
S16 |
S17 |
S18 |
S19 |
S20 |
S21 |
S22 |
S23 |
S24 |
S25 |
S26 |
S27 |
S28 |
S29 |
S30 |
S31 |
S32 |
S33 |
S34 |
|
Zacco platypus
|
4 |
6 |
10 |
3 |
39 |
49 |
57 |
51 |
48 |
81 |
106 |
119 |
97 |
94 |
117 |
45 |
68 |
74 |
110 |
146 |
132 |
105 |
118 |
137 |
282 |
134 |
127 |
35 |
38 |
26 |
29 |
39 |
47 |
61 |
2634 |
35.4 |
100 |
|
Zacco koreanus
|
65 |
65 |
75 |
77 |
38 |
37 |
47 |
32 |
60 |
20 |
27 |
43 |
63 |
86 |
16 |
5 |
2 |
|
22 |
2 |
18 |
46 |
10 |
8 |
6 |
|
7 |
|
|
|
|
|
|
|
877 |
11.8 |
73.5 |
|
Carassius. auratus
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
14 |
142 |
79 |
31 |
2 |
28 |
5 |
1 |
15 |
1 |
6 |
14 |
82 |
104 |
98 |
23 |
|
|
|
|
645 |
8.7 |
47.1 |
|
Acheilognathus lanceolatus
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
95 |
110 |
35 |
54 |
77 |
21 |
38 |
17 |
18 |
12 |
21 |
41 |
24 |
26 |
1 |
|
|
2 |
2 |
16 |
612 |
8.2 |
55.9 |
|
Iksookimia koreensis
|
4 |
|
3 |
2 |
6 |
17 |
22 |
22 |
5 |
40 |
49 |
32 |
17 |
25 |
22 |
|
|
10 |
2 |
|
1 |
11 |
3 |
1 |
3 |
1 |
|
|
|
|
|
5 |
|
6 |
309 |
4.2 |
70.6 |
|
Rhinogobius brunneus
|
|
|
|
|
|
|
|
|
|
|
1 |
1 |
3 |
|
6 |
2 |
11 |
6 |
3 |
6 |
9 |
21 |
23 |
31 |
22 |
25 |
25 |
12 |
6 |
9 |
11 |
18 |
16 |
23 |
290 |
3.9 |
67.6 |
|
Pseudogobio esocinus
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
13 |
3 |
1 |
2 |
7 |
1 |
2 |
6 |
17 |
16 |
70 |
15 |
7 |
14 |
2 |
4 |
23 |
16 |
16 |
19 |
254 |
3.4 |
58.8 |
|
Pungtungia herzi
|
|
2 |
5 |
1 |
|
3 |
7 |
1 |
9 |
9 |
8 |
11 |
8 |
5 |
8 |
11 |
27 |
|
18 |
6 |
12 |
13 |
17 |
16 |
16 |
7 |
6 |
16 |
5 |
|
|
2 |
|
|
249 |
3.4 |
79.4 |
|
Microphysogobio yaluensis
|
|
|
|
|
|
|
|
1 |
|
7 |
|
1 |
|
14 |
25 |
34 |
2 |
|
|
10 |
9 |
22 |
5 |
25 |
9 |
33 |
14 |
|
|
|
|
7 |
13 |
15 |
246 |
3.3 |
52.9 |
|
Micropterus salmoides
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3 |
2 |
38 |
30 |
53 |
59 |
6 |
25 |
6 |
1 |
223 |
3.0 |
29.4 |
|
Rhynchocypris oxycephalus
|
72 |
36 |
15 |
14 |
19 |
8 |
4 |
13 |
2 |
3 |
3 |
2 |
4 |
8 |
7 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
210 |
2.8 |
44.1 |
|
Acheilognathus rhombeus
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
12 |
5 |
16 |
7 |
1 |
|
|
2 |
|
10 |
26 |
11 |
25 |
68 |
|
|
|
|
|
183 |
2.5 |
32.4 |
|
Hemibarbus longirostris
|
|
|
|
|
|
|
|
1 |
|
|
3 |
|
3 |
2 |
4 |
|
|
6 |
1 |
|
|
|
18 |
|
2 |
4 |
2 |
15 |
1 |
6 |
|
27 |
4 |
1 |
100 |
1.3 |
50.0 |
|
Obontobutis interrupta
|
|
2 |
|
2 |
|
|
|
2 |
2 |
1 |
4 |
2 |
7 |
3 |
3 |
5 |
7 |
14 |
7 |
7 |
|
4 |
9 |
3 |
1 |
|
|
|
1 |
1 |
2 |
1 |
3 |
6 |
99 |
1.3 |
73.5 |
|
Cyprinus carpio
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
48 |
35 |
|
|
|
1 |
|
1 |
85 |
1.1 |
11.8 |
|
Others (RA less then 1%) |
1 |
|
4 |
4 |
4 |
1 |
|
6 |
1 |
1 |
1 |
2 |
15 |
3 |
11 |
17 |
21 |
20 |
24 |
15 |
17 |
4 |
28 |
29 |
32 |
61 |
2 |
2 |
5 |
28 |
6 |
8 |
9 |
38 |
420 |
5.6 |
- |
|
No. of species |
5 |
5 |
7 |
7 |
6 |
6 |
5 |
9 |
7 |
8 |
9 |
9 |
9 |
10 |
16 |
19 |
16 |
12 |
18 |
13 |
15 |
14 |
20 |
18 |
20 |
18 |
14 |
12 |
13 |
9 |
7 |
16 |
11 |
16 |
5 |
- |
- |
|
No. of individuals |
146 |
111 |
112 |
103 |
106 |
115 |
137 |
129 |
127 |
162 |
202 |
213 |
217 |
242 |
341 |
386 |
258 |
233 |
280 |
243 |
243 |
250 |
283 |
279 |
483 |
363 |
393 |
314 |
278 |
156 |
77 |
151 |
116 |
187 |
146 |
100 |
- |
Table 3.
Indicator value analysis based on fish assemblages of each group.
|
Groups |
Species name |
IndVal |
p-value |
Frequency (sites) |
|
A (Rhynchocypris community) |
Rhynchocypris oxycephalus
|
0.840 |
< 0.001 |
15 |
|
Zacco koreanus
|
0.566 |
< 0.001 |
25 |
|
Misgurnus anguillicaudatus
|
0.419 |
0.034 |
8 |
|
B (Iksookimia community) |
Iksookimia koreensis
|
0.777 |
< 0.001 |
24 |
|
C (Pseudogobio community) |
Pseudogobio esocinus
|
0.745 |
< 0.001 |
20 |
|
Rhodeus ocellatus
|
0.538 |
0.010 |
7 |
|
Rhinogobius brunneus
|
0.537 |
0.003 |
23 |
|
Pseudorabora parva
|
0.494 |
0.017 |
13 |
|
Zacco platypus
|
0.441 |
0.006 |
34 |
|
D (Pseudogobio community) |
Carassius auratus
|
0.907 |
< 0.001 |
16 |
|
Acheilognathus rhombeus
|
0.674 |
0.003 |
11 |
|
Micropterus salmoides
|
0.600 |
0.005 |
10 |
Table 4.
The FAI score of each group and its metrics based on fish assemblages (M1: Number of native species, M2: Number of riffle-benthic species, M3: Number of sensitive species, M4: Proportion of tolerant species, M5: Proportion of omnivores, M6: Proportion of native insectivores, M7: Number of native individuals, M8: Proportion of anomalies)
|
Groups |
M1 |
M2 |
M3 |
M4 |
M5 |
M6 |
M7 |
M8 |
FAI |
|
A |
12.5 |
1.6 |
7.8 |
12.5 |
12.5 |
12.5 |
12.5 |
12.5 |
84.4 |
|
B |
11.9 |
5.1 |
5.7 |
12.5 |
6.3 |
10.8 |
12.5 |
12.5 |
77.3 |
|
C |
11.5 |
4.8 |
0.5 |
12.5 |
2.9 |
4.8 |
11.5 |
12.5 |
61.1 |
|
D |
10.4 |
2.1 |
0.0 |
7.3 |
1.0 |
1.0 |
11.5 |
12.5 |
45.8 |
References
1. A. J. Reid, A. K. Carlson, I. F. Creed, E. J. Eliason, P. A. Gell, P. T. J. Johnson, K. A. Kidd, T. J. MacCormack, J. D. Olden, S. J. Ormerod, J. P. Smol, W. W. Taylor, K. Tockner, J. C. Vermaire, D. Dudgeon, S. J. Cooke, Emerging threats and persistent conservation challenges for freshwater biodiversity, Biol. Rev., 94, 849-873(2019).   2. B. Grizzetti, C. Liquete, A. Pistocchi, O. Vigiak, G. Zulian, F. Bouraoui, A. De Roo, A.C. Cardoso, Relationship between ecological condition and ecosystem services in European rivers, lakes and coastal waters, Sci. Total Environ., 671, 452-465(2019). 3. C. M. Pringle, Hydrologic connectivity and the management of biological reserves: a global perspective, Ecol. Appl., 11(4), 981-998(2001). 4. J. Xia, Y. Gao, Q. T. Zuo, X. J. Liu, Q. M. Chen, M. Dou, Characteristics of interconnected river system and its ecological effects on water environment, Prog. Geogr., 31(1), 26-31(2012).
5. R. L. Vannote, G. W. Minshall, K. W. Cummins, J. R. Sedell, C. E. Cushing, The River Continuum Concept, Can. J. Fish. Aquat. Sci., 37, 130-137(1980). 6. S. T. A. Pickett, M. L. Cadenasso, E. J. Rosi-Marshall, K. T. Belt, P. M. Groffman, J. M. Grove, E. G. Irwin, S. S. Kaushal, S. L. LaDeau, C. H. Nilon, C. M. Swan, P. S. Warren, Dynamic heterogeneity: a framework to promote ecological integration and hypothesis generation in urban systems, Urban Ecosyst., 20, 1-14(2017). 7. J. A. Roebuck Jr, M. Seidel, T. Dittmar, R. Jaffe, Controls of land use and the river continuum concept on dissolved organic matter composition in an anthropogenically disturbed subtropical watershed, Environ. Sci. Technol., 54(1), 195-206(2019). 8. M.D. Delong, M.A. Brusven, Macroinvertebrate community structure along the longitudinal gradient of an agriculturally impacted stream, Environ. Manag., 22(3), 445-457(1998). 9. A. Vila-Gispert, E. García-Berthou, R. Moreno-Amich, Fish zonation in a Mediterranean stream: Effects of human disturbances, Aquat. Sci., 64, 163-170(2002). 10. M. Dynesius, C. Nilsson, Fragmentation and flow regulation of river systems in the northern third of the world, Science., 266, 753-762(1994). 11. P. Roni, K. Hanson, T. Beechie, Global review of the physical and biological effectiveness of stream habitat rehabilitation techniques, N. Am. J. Fish. Manag., 28, 856-890(2008). 12. C. S. Park, K. G. An, Fish passage assessments in the fishway of Juksan weir constructed in the downstream area of Youngsan-River watershed, J. Environ. Sci. Int., 23(8), 1513-1522(2014). 13. M. C. Lucas, E. Baras, Migration of freshwater fishes. Blackwell Science, Oxford, UK(2001).
14. J. E. Wofford, R. E. Gresswell, M. A. Banks, Influence of barriers to movement on within‐watershed genetic variation of coastal cutthroat trout, Ecol. Appl., 15(2), 628-637(2005). 15. N. Poulet, Impact of weirs on fish communities in a Piedmont stream, River Res. Applic., 23, 1038-1047(2007). 16. M. Larinier, Environmental issues, dams and fish migration, FAO fisheries technical paper., 419, 45-89(2001).
17. L. Carpenter-Bundhoo, G. L. Butler, N. R. Bond, S. E. Bunn, I. V. Reinfelds, M. J. Kennard, Effects of a low-head weir on multi-scaled movement and behavior of three riverine fish species, Sci. Rep., 10, 6817(2020; https://doi.org/10.1038/s41598-020-63005-8. 18. D. H. Yeom, S. A. Lee, G. S. Kang, J. Seo, S. K. Lee, Stressor identification and health assessment of fish exposed to wastewater effluents in Miho Stream, South Korea. Chemosphere., 67(11), 2282-2292(2007).
19. K. G. An, S. J. Lee, Conservation and Management in Korea Using Fish Multi-Metric Model, Korean J. Ecol. Environ., 51(1), 86-95(2018).
20. D. H. Yeom, K. H. Chung, Y. H. Kim, S. M. Adams, Ecological health and causal assessment of fish communities experiencing multiple stressors in Gap Stream, South Korea, Toxicol. Environ. Health. Sci., 1, 97-108(2009; https://doi.org/10.1007/BF03216470. 21. H. K. Park, J. K. Choi, K. H. Won, H. G. Lee, Study on the evaluation of ecological health by using fish communities in the Wonju stream, Korea. Korean J. Environ. Biol., 35(4), 684-693(2017).
22. J. K. Choi, H. G. Byeon, H. G. Seok, Studies on the dynamics of fish community in Wonju stream, Korean J. Limnol., 33(3), 274-281(2000).
23. J. H. Han, D. G. Ko, B. J. Lim, J. H. Park, K. G. An, Summer patterns and diel variations of fish movements using fish trap sampling technique in the Juksan weir, J. Environ. Impact Assess., 21(6), 879-891(2012).
24. T. G. Northcote, Migratory behaviour of fish and its significance to movement through riverine fish passage facilities. Fish Migration and Fish BypassesM. Jungwirth, S. Schmutz, S. Weiss(Eds.), Fishing News Books, Oxford, England, pp. 3-18(1998).
25. P. S. Kemp, J. R. O’Hanley, Procedures for evaluating and prioritising the removal of fish passage barriers: a synthesis, Fish. Manag. Ecol., 17(4), 297-322(2010). 26. J. Y. Kim, J. G. Kim, D. Y. Bae, H. J. Kim, J. E. Kim, H. S. Lee, J. Y. Lim, K. G. An, International and domestic research trends in longitudinal connectivity evaluations of aquatic ecosystems, and the applicability analysis of fish-based models, Korean J. Environ. Biol., 38(4), 634-649(2020). 27. MOE/NIER, Freshwater ecosystem survey and assessment manual Ministry of Environment/National Institute of Environmental Research, Incheon, South Korea(2020).
28. Ohio EPA, Biological Criteria for the Protection of Aquatic Life Ohio Environmental Protection Agency, Columbus, OH, USA(1987).
29. M. T. Barbour, J. Gerritsen, B. Snyder, J. Stribling, Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, benthic macroinvertebrates, and fish, USEPA., 339, 1-340(1999).
30. J. R. Karr, Assessment of Biotic Integrity Using Fish Communities, Fisheries., 6, 21-27(1981). 31. MOE/NIER, Biomonitoring survey and assessment manual Ministry of Environment/National Institute of Environmental Research, Incheon, South Korea(2017).
32. R Core Team, R: A language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria(2023).
33. J. Oksanen, F. G. Blanchet, R. Kindt, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, P. R. Minchin, R. B. O’Hara, G. L. Simpson, P. Solymos, M. H. H. Stevens, E. Szoeces, H. Wagner, Package ‘vegan’, https://cran.r-hub.io/web/packages/vegan/vegan.pdf, (2019) 34. D. Borcard, F. Gillet, P. Legendre, Numerical ecology with R Springer, New York, USA(2018).
35. J. H. Ward, Hierarchical grouping to optimize an objective function, J. Am. Stat. Assoc., 58, 236-244(1963). 36. J. B. Kruskal, Nonmetric multidimensional scaling: a numerical method, Psychometrika., 29(2), 115-129(1964). 37. J. R. Bray, J. T. Curtis, An ordination of the upland forest communities of Southern Wisconsin, Ecol. Monogr., 27(4), 325-349(1957). 39. M. Dufrêne, P. Legendre, Species assemblages and indicator species: the need for a flexible asymmetrical approach, Ecol. Monogr., 67(3), 345-366(1997). 40. H. Kang, D. Im, J. W. Hur, K. H. Kim, Instream of Habitat Suitability Index of Fish Species in the Geum River Watershed, J. Korean Soc. Civil Eng., 31(2B), 193-203(2011).
41. W. K. Moon, J. H. Han, K. G. An, Fish fauna and community analysis in Heuck Stream watershed, Korean J. Limnol., 43, 69-81(2010).
42. K. Lange, C. R. Townsend, R. Gabrielsson, P. C. Chanut, C. D. Matthaei, Responses of stream fish populations to farming intensity and water abstraction in an agricultural catchment, Freshw. Biol., 59(2), 286-299(2014).
|
|










